
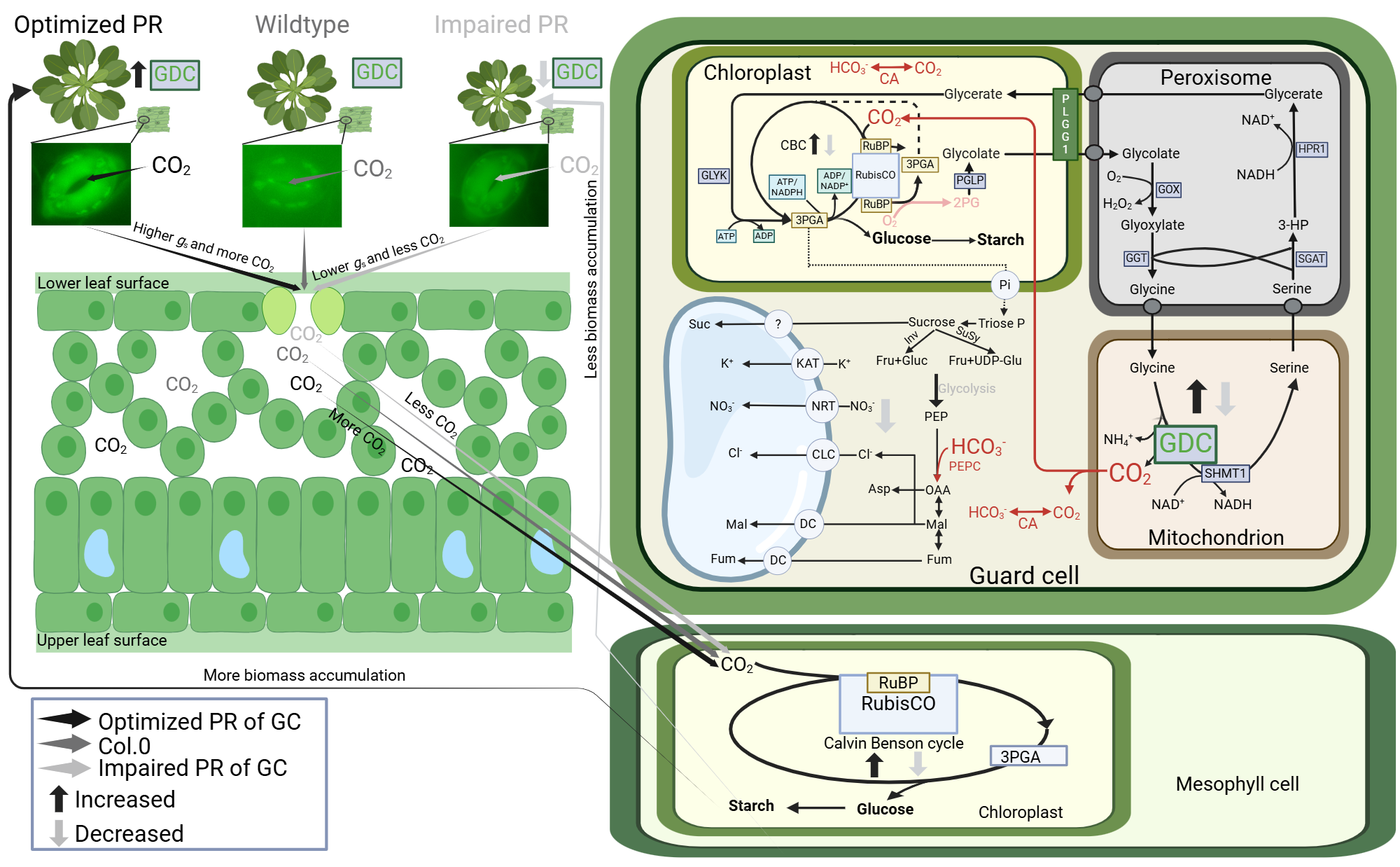

Stomata physiologically respond to changes in external CO2 availability, in turn affecting internal CO2 (Ci). High CO2 leads to stomatal closure, whilst low CO2 leads to stomatal opening. We hypothesize that changes in Ci, and the resulting movements, can be mimicked through our transgenic intervention, assumed to alter guard cell photorespiratory metabolism (i.e. GDC-H protein expression). At one hand, low external CO2 stimulates photorespiratory metabolism (more Rubisco oxygenation), which is similar in the GDC-H overexpression plants, and prompts stomata to be more open (low Ci). On the other hand, high external CO2 suppresses photorespiratory metabolism (less Rubisco oxygenation), which is comparable with our guard cell GDC-H antisense lines, causing stomata to be more closed (high Ci). Mechanistically, improved or lower (impaired) GC photorespiratory metabolism is hypothesized to alleviated or enhanced negative metabolic feedback onto the CB-cycle, improving or hindering RuBP regeneration. Improved CB-cycle performance lowers Ci and makes GC a stronger CO2 sink, enhances stomatal opening (higher gs), to ultimately provide more CO2 for mesophyll photosynthesis. Higher photosynthesis stimulated carbohydrate biosynthesis and plant growth. Impaired or repressed CB-cycle performance lowers CO2 requirements, leads to less open stomata (lower gs) and decreases mesophyll CO2. Reduced mesophyll photosynthesis provides less carbon for plant growth. However, given experimental evidence revealed only low Rubisco amounts and CB-cycle activity in GC, this hypothesis is only partially sufficient to explain the physiological responses. Thus, we further assume that manipulation of GC GDC activity (glycine decarboxylation) through, increased and suppressed GDC-H protein expression, has a direct impact on the GC Ci, too. More GDC activity (in overexpression lines, increased glycine decarboxylation) leads to increased, whilst lower GDC activity (in antisense lines, reduced glycine decarboxylation) leads to reduced GC Ci. Changes in GC Ci supports (or reduces) photosynthesis for increased (or decreased) GC starch production, providing more (or less) energy supply for stomatal movements. Changes in CO2 are likely to affect PEPC mediated GC photosynthesis. It seems also reasonable to consider an impact on GC nitrate and H2O2 amounts, as a consequence of altered GDC activity and photorespiration, as additional factors leading to changes in GC metabolism and stomatal movements. The figure was created with BioRender (https://www.biorender.com/[https://BioRender.com/b29k163]).Abbreviations: enzymes; CA - carbonic anhydrase; Inv - invertase; PEPc - phosphoenolpyruvate carboxylase; PEPk - phosphoenolpyruvate carboxykinase; RubisCO - ribulose-1,5-biphosphate carboxylase/oxygenase; SuSy - sucrose synthase. Metabolites; 3PGA - 3 phosphoglycerate; Asp - aspartate; Cl−- chloride; Fru - fructose; Fum - fumarate; Glu - glutamate; Gluc - glucose; Isoc - isocitrate; K+- potassium; Mal - malate; NO3− - nitrate; OAA - oxaloacetate; PEP, phosphoenolpyruvate; Suc - sucrose; UDP-glu - uridine diphosphate glucose. Transporters; CLC - chloride channel; DC - dicarboxylate carrier; NRT - nitrate transporter; OC - putative oxaloacetate carrier.
